番茄基因组逆转座子的鉴定与演化分析 
 作者
作者  通讯作者
通讯作者
《分子植物育种》网络版, 2013 年, 第 11 卷, 第 25 篇 doi: 10.5376/mpb.cn.2013.11.0025
收稿日期: 2013年06月24日 接受日期: 2013年06月26日 发表日期: 2013年08月08日
引用格式(中文):
许莹修, 2013, 番茄基因组逆转座子的鉴定与演化分析, 分子植物育种(online), 11(25): 1181-1189 (doi: 10.5376/mpb.cn.2013.11.0025)
引用格式(英文):
Xu Y.X., 2013, Genome-scale Identification and Evolutionary Analysis of LTR Retrotransposons in Tomato, Fenzi Zhiwu Yuzhong (online) (Molecular Plant Breeding), 11(25): 1181-1189 (doi: 10.5376/mpb.cn.2013.11.0025)
逆转座子是真核生物特别是高等植物基因组的重要组分,对其演化起到关键作用。为了解析逆转座子在番茄基因组中的分布及其在基因组进化中的作用,采用结构分析和同源比较方法从全基因组水平鉴定了栽培番茄的LTR逆转座子,得到15 036个逆转座元件,共86.01 Mb,占测序序列的11.02%。将这些元件分为330个家族,其中copia类的182个家族,gypsy类80个。完整转座元件的转座时间在2.5 MYA 时间处于高峰,比大豆、玉米等逆转座元件整体古老。比较栽培番茄与野生醋栗番茄中完整转座子的时空分布发现栽培番茄(Solanum lycopersicum)与野生番茄(Solanum pimpinellifolium)在最近的时间段内(25万年以内)产生了较明显的分化。有930 个逆转座元件插入到基因序列附近或内部,可能在物种演化中起到了重要作用。
DNA重复序列是构成真核生物基因组的主要组分,其中移动元件(mobile elements)占有很大比例。逆转座子(retrontransposon)是一类通过“复制-粘贴”方式转座的移动元件,在真核生物中特别是许多植物基因组中普遍存在(Finnegan, 1989; Kumar and Bennetzen, 1999; Sabot and Schulman, 2006; SanMiguel et al., 1996; Wicker et al., 2007)。其中LTR逆转座子两端有正向长末端重复序列(long terminal repeat, LTR),中间是一个或几个开放阅读框(ORF),编码转座所需的各个蛋白组分。依据编码各组分的序列排列顺序的不同和序列同源性LTR逆转座子大体分为Ty1-copia和Ty3-gypsy两类(Havecker et al., 2004)。LTR逆转座子在真核生物中含有大量拷贝,是构成基因组尤其是高等植物基因组序列的主要组分(Baucom et al., 2009; Du et al., 2010; International Rice Genome Sequencing Project, 2005; Ma et al., 2004; Schnable et al., 2009)。碱基突变和序列重组是基因组变异的主要来源,而逆转座子通过转座、删除机制可以产生大量的序列变化,带来丰富的遗传信息,因此分析研究逆转座子的分布和变化是了解基因组进化发展和生物演化的重要手段。
番茄是重要的经济作物和研究果实发育的模式植物,许多研究工作都尝试在番茄基因组中进行逆转座子的分析鉴定(Rogers and Pauls, 2000; Su and Brown, 1997; Yañez et al., 1998),而且番茄中已开发出基于逆转座子序列的多态性较高的分子标记系统(Tam et al., 2005)。逆转座子在栽培番茄基因组演化和物种形成中起到了重要的作用,包括其复制和转座对功能基因的影响。例如逆转座子Rider介导的一个SUN基因的重复使得其表达水平升高,从而改变了番茄果实的形态(Xiao et al., 2008)。Rider是一个近期活跃的家族,部分成员已经被分离鉴定,研究表明Rider 可能是在1~6百万年(MYA)之间,通过水平转移方式进入番茄基因组的(Cheng et al., 2009; Jiang et al., 2009)。再比如一个copia逆转座子插入到一个高活性的酯酶基因附近,增加了它的表达量,降低了其底物酯类的水平,从而影响了番茄果实的味道(Goulet et al., 2012)。番茄全基因组序列测定的完成,为全面分析其LTR逆转座子提供了条件,测序结果也表明番茄基因组中含有大量逆转座子序列(The Tomato Genome Consortium, 2012)。
本研究基于番茄全基因组测序结果,利用生物信息学方法从整个基因组水平鉴定逆转座子,对其组成、分布特点和种类等进行分析,对其在栽培和野生醋栗番茄之间的差异进行了比较,并探讨了逆转座子在番茄物种演化和基因功能进化中的作用。
1结果和分析
1.1逆转座子及相关组分的分离鉴定和分类
用LTR_STRUC 程序对整个基因组进行分析,分离得到1 825个结构完整的候选转座子。人工检测确定1 410个结构完整清晰的元件作为进一步研究的基础。用perl 编写程序,在番茄基因组中分析上一步得到的转座子同源序列,得到676个完整元件,9 408个非完整元件及3 542个solo元件,共计15 036个(86.08 Mb),占基因组测序序列的11.02% (表1)。另外检测到284.04 Mb的同源DNA片段,占基因组序列的36.19%。完整元件共2 086个,序列总长度15.80 Mb,单个长度介于943 bp和23 668 bp之间,平均7 605 bp。
|
|

对2 086个人工确认的完整转座子的LTR 序列进行聚类分析,依据其遗传聚类和相似程度,共得到330个家族,其他元件根据其最初鉴定所依据的完整LTR序列划分到相应的家族,其中182个copia 类,80个gypsy 类,其他68 个未能鉴定(not determined, ND)(表2)。有29个家族含有100个以上元件(表3),其中3个家族SL_RT_F94、SL_RT_F324和SL_RT_F322含有超过1 000个元件,均是gypsy 类型。
|
|

|
|

1.2转座子的转座时间和分布
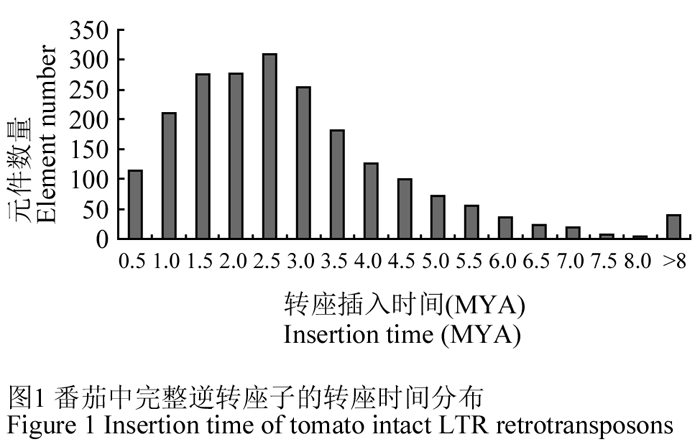
逆转座子转座发生的时间是重要的信息,而且可以依据其两侧LTR序列的差异判断突变的比率从而确定转座发生至今的大概时间(SanMiguel et al., 1998)。比较分析2 086个完整转座子的LTR序列并计算突变率,确定了它们的转座年龄,结果表明在不同的时间段转座发生的频率是不同的。转座高峰时期在2.5 MYA 时间左右,表明在这个时间段番茄基因组中逆转座子活性较强(图1)。番茄逆转座子转座高峰时期与其他已知的高等植物有所不同,在接近现代时间段内转座事件逐渐减少,而如大豆、玉米等基因组中转座事件却随时间接近而逐渐增加(Baucom et al., 2009; Du et al., 2010)。
|
|
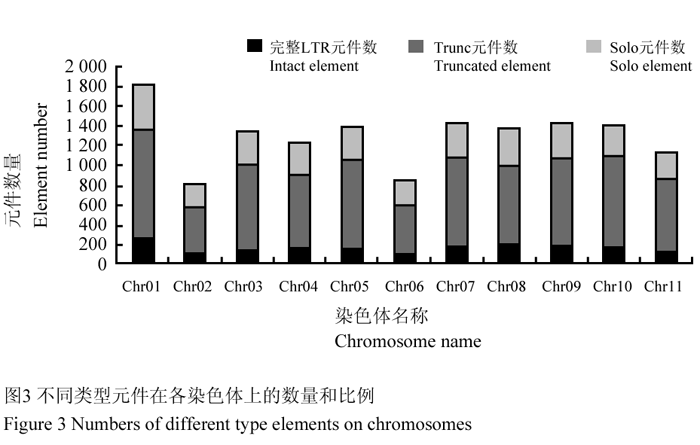
我们统计了不同染色体上逆转座元件的数量及其与各染色体测序长度的关系,结果表明二者基本成正比(图2),不同种类逆转座元件在各染色体上的分布也大体均衡,各染色体上都是非完整元件所占比例最大,完整元件较少(图3)。
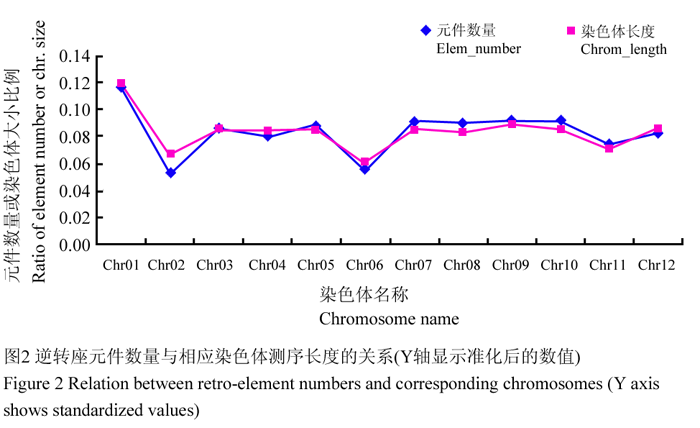 图2 逆转座元件数量与相应染色体测序长度的关系(Y轴显示准化后的数值) Figure 2 Relation between retro-element numbers and corresponding chromosomes (Y axis shows standardized values) |
|
|
1.3完整逆转座子元件在栽培和野生番茄中的差异分析
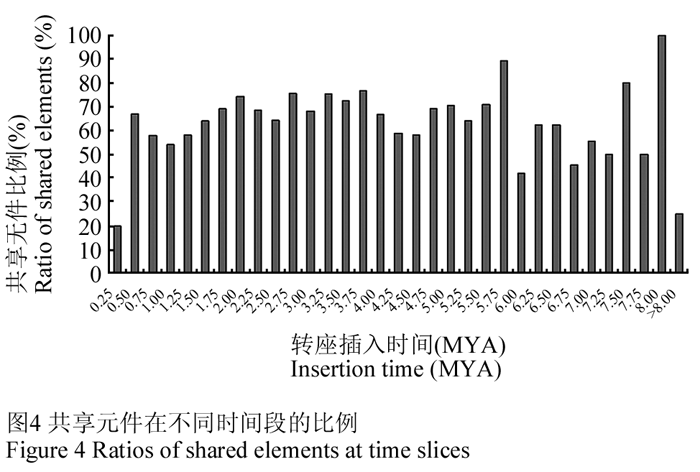
除去单个碱基的自然突变之外,逆转座子还可以通过序列重组过程不断产生大量插入、缺失及异位重组等变异(Girard and Freeling, 1999; Morgante et al., 2007)。逆转座子的转座和消除机制以及突变等过程表明它们可以在较短时间内改变基因组序列组成并影响基因功能,而且这种快速的变化使得在不同物种之间逆转座子很少存在较高的序列同源性。但是在近缘物种中比较逆转座子的差异可以为研究物种分化、植物驯化及品种差异等提供丰富的信息。在番茄栽培种和野生种分化后发生的逆转座子转座事件必然在两个基因组中产生差异,依据发生转座的时间可以判断差异产生是在物种分化以前或以后,反之也可以推测物种分化的大体时间。我们分析了栽培番茄中完整转座子在野生番茄基因组中的相应分布,结果发现1 341 个元件在两个基因组中是相同的(shared),但是栽培番茄中另外鉴定到733 个转座元件在野生番茄中并不存在,说明是在物种分化后形成或者是在野生番茄基因组进化过程中被删除(表1)。依据不同时间段计算不同转座元件的比例的话,可以看出在最近的0.25 MYA时间段内shared 元件的比例明显高于其它所有时间段,表明在最近的25 万年里,栽培番茄基因组与野生番茄基因组发生了明显的分化 (图4)。
|
|
1.4 插入编码基因内部或附近的逆转座元件
逆转座子对基因组进化的影响是多方面的。从功能角度来讲基因组的演化包括基因表达变化(包括调控序列的碱基变化和表观修饰)、编码序列的改变和可变剪切,而逆转座子的活动对这几个方面都可以产生影响(Hua-Van et al., 2011)。逆转座子也可以通过插入或者删除影响基因组的大小以及染色体结构。插入基因序列附近甚至内部的逆转座子元件对于基因组的演化具有较直接的影响,可能在物种进化过程中起到了重要作用。我们比较了栽培番茄中所有逆转座元件与基因的相对位置,结果发现有930 个元件位于基因序列附近(与基因序列5 '端距离小于1 000 bp 或位于基因内部)(表4)。
|
|

2讨论
2.1番茄基因组中逆转座子的数量和分布
我们用结构分析和同源比对方法在番茄基因组中鉴定到15 036 个逆转座元件,总共86.1 Mb,占测序序列总长度(759.9 Mb)的11.3%。这些元件不包括零散的难于精确识别的逆转座子序列片断。其中完整元件有2 068 个,只有总数的13.7%,说明大多数元件的结构已经随着时间的流逝通过突变和各种消除机制被破坏。从转座时间分布来看也符合这种推测。番茄完整转座子的转座插入时间峰值在2.5 MYA 左右,在这之后就迅速下降。而在大豆基因组中逆转座子随时间的接近数量不断增加,转座插入时间峰值在0~0.5 MYA (Du et al., 2010),玉米中LTR逆转座子也表现出相似的分布模式(Baucom et al., 2009),说明番茄中大多数逆转座元件相对古老,发生突变和结构变化的比例会更高。
逆转座元件变异迅速,种类繁多,并没有统一的分类系统。依据pol基因编码产物的排列顺序和序列同源性LTR转座子可分为copia 和gypsy 两个主要类型。我们依据前人的一些标准(Wicker et al., 2007)用同源分类方法将番茄中逆转座子分为330 个家族,其中鉴定到copia类(182个)远多于gypsy (80个),但是从元件数量上来看copia 类(5 314个)却少于gypsy (8 978个),并且含有1 000个以上元件的三个家族都是gypsy 类型。因此,copia 逆转座元件的多态性较高,而gypsy 在进化中相对保守。
研究表明逆转座元件的转座插入位置序列并没有特异性,是随机的过程。但是逆转座子在染色体上的分布有一定的差异,通常在基因较多的区域,逆转座元件相对较少,而基因密度小的区域逆转座元件相对较多(Baucom et al., 2009)。这种不均匀可能是由于逆转座子在转座时对特定区域有一定选择性或者是由于逆转座子的消除机制更多地作用于常染色质区,也可能是不同区域所面对的选择压力差异的反映。番茄中逆转座元件在12 条染色体上分布大体均匀,数量基本与染色体序列长度成正比,表明对于不同染色体来讲,逆转座子的转座并没有选择性。
2.2番茄中逆转座子的转座时间及种间差异比较
逆转座子的转座机制决定了它们在转座后两端的LTR序列是完全一致的,因此比较完整转座子两侧LTR序列就可以依据其碱基突变率计算其年龄。同时由于逆转座子的转座插入是随机的过程并且转座事件在物种进化的各个时期都在不断进行,因而可以为研究物种及其基因组的分化和演化提供丰富的信息。由地理隔离发展到生殖隔离,即种群间不存在基因交流是物种形成的条件和标志,许多作物栽培种群的形成是人为阻断基因交流并产生生殖隔离的过程。多位点突变率计算表明栽培番茄与醋栗番茄在1.4 MYA 发生分化(Nesbitt and Tanksley, 2002),同时研究表也明栽培番茄的起源和驯化伴随着野生番茄基因的不断渗入,而樱桃番茄 S. lycopersicum var. cerasiforme (S. l. cerasiforme)被认为是野生番茄向栽培番茄发展的一个过渡种(Nesbitt and Tanksley, 2002; Ranc et al., 2008)。逆转座子的copy-paste 转座模式决定了它的每一个转座事件都会在基因组进化中留下特异的标记,又由于可以借助两端LTR 的差异解析其自身存在的时间,因此含有大量的进化信息。我们分析栽培番茄和醋栗番茄之间LTR逆转座子的等位性分布发现在大部分时间段内都存在很大比例在两个基因组中分布相同(shared)的LTR逆转座元件,表明的确野生番茄和栽培番茄的基因组序列存在遗传物质交流。在物种分化后期基因组必然是独立演化的,因此这个时间以后发生的逆转座子的复制和删除应该是物种特异的,而在这个时间点之前发生的转座子复制在两个物种中应该大部分保持相同的分布。栽培和野生醋栗番茄中0.25 MYA 以上的时间段内相同分布的逆转座元件比例均在50% 以上,但是在0.25 MYA 以内双方共有的LTR 元件大大低于其他时间段(约20%),表明在近期栽培种与野生种产生了较明显的分化,是栽培番茄物种形成的阶段,明显晚于已知的分化时间。
2.3逆转座子对功能基因进化的影响
逆转座子的插入位点序列是随机的,而且在真核生物特别是高等植物中存在大量转座事件,因此从理论上来说逆转座子可以插入基因附近影响其功能,甚至进入基因内部破坏其结构。很多研究已经表明逆转座子可以影响基因的表达,甚至通过自身含有的转座元件直接调控基因表达(Roman et al., 2008)。基因的启动子包括核心启动子区域和调控区域,总长度可以在2 000 bp以上。我们发现有930个元件位于基因上游1 000 bp 以内或者位于基因内部,这些元件在转座过程中很可能会影响基因的表达调控及生物学功能。除直接插入到基因内部的元件以外,还有22个元件位于基因上游35 bp以内,可以肯定这些元件在转座过程中至少改变了原始基因的结构或表达水平。
3材料和方法
3.1材料
栽培番茄(Solanum lycopersicum)和野生醋栗番茄(Solanum pimpinellifolium)全基因组序列S_lycopersicum_chromosomes.2.40和S_lycopersicum_chromosomes.2.40.fa来源于ftp://ftp.sgn.cornell.edu/。
3.2方法
3.2.1 LTR-逆转座子的鉴定
用结构分析和同源比对的方法鉴定反转座子元件。首先用LTR_STRUC程序(Mccarthy and Mcdonald, 2003)寻找完整结构的逆转座子,然后利用其LTR 序列进行同源比对,并应用perl语言编程从中寻找非完整元件(truncated elements)和solo 元件(Ma et al., 2004)。
3.2.2逆转座子的分类
逆转座子家族的分类方法依据前人的标准修改后进行(Wicker et al., 2007)。对完整逆转座子的LTR序列用clustalW 程序进行聚类分析,计算各个元件的遗传距离,差异小于0.2 的转座元件划分为同一家族。其它同源分析方法得到的转座元件依据其匹配的相应完整元件确定所属家族。转座子家族种类的确定(copia 或者gypsy)利用最近时间段内发生转座插入的元件,用NCBI的RPS-BLAST程序分析是否存在与copia 或gypsy 转座子同源的反转录酶蛋白序列(Marchler-Bauer et al., 2009; Marchler-Bauer and Bryant, 2004; Marchler-Bauer et al., 2011; Xiong and Eickbush, 1990)。
3.2.3转座插入时间鉴定
通过比较完整转座子两端的LTR 序列来确定转座子的插入时间。首先用MUSCLE程序比对转座子两端的LTR 序列(Edgar, 2004),计算比对碱基的总数和碱基突变的数目,计算得到碱基替换率(r)。遗传距离(K)用Jukes–Cantor方法(Kimura and Ohta, 1972)进行修正。以平均每年每个位点1.3×10-8 碱基替换数作为计算转座插入年龄的依据(Ma and Jackson, 2006)。最后转座子的插入时间(T)用公式T = K/2r 计算得到。
3.2.4栽培番茄逆转座子在野生醋栗番茄的等位鉴定
依据已有的方法完成(Tian et al., 2009)。完整转座元件的5'和3'端侧翼序列(25 bp)结合转座子本身边界序列(25 bp)用cross_match 在野生醋栗番茄中做序列比对,5'或者3'结合序列在野生番茄基因组中有良好匹配结果的认为该元件在两个物种中是都存在的(shared),如果5'和3'序列均不存在匹配结果,则该元件在栽培番茄中是特异的(unshared)。
作者贡献
许莹修是本研究的构思者及负责人,实验设计,数据分析,论文写作与修改。
致谢
本研究得到江苏省农业科技自主创新基金(CX(12)5037)资助。
参考文献
Baucom R.S., Estill J.C., Chaparro C., Upshaw N., Jogi A., Deragon J.M., Westerman R.P., Sanmiguel P.J., and Bennetzen J.L., 2009, Exceptional diversity, non-random distribution, and rapid evolution of retroelements in the B73 maize genome, PLoS Genet, 5(11): e1000732
http://dx.doi.org/10.1371/journal.pgen.1000732
PMid:19936065 PMCid:PMC2774510
Cheng X.D., Zhang D.F., Cheng Z.K., Keller B., and Ling H.Q., 2009, A new family of Ty1-copia-like retrotransposons originated in the tomato genome by a recent horizontal transfer event, Genetics, 181(4): 1183-1193
http://dx.doi.org/10.1534/genetics.108.099150
PMid:19153256 PMCid:PMC2666490
Du J.C., Tian Z.X., Hans C.S., Laten H.M., Cannon S.B., Jackson S.A., Shoemaker R.C., and Ma J.X., 2010, Evolutionary conservation, diversity and specificity of LTR-retrotransposons in flowering plants: insights from genome-wide analysis and multi-specific comparison, Plant J., 63(4): 584-598
http://dx.doi.org/10.1111/j.1365-313X.2010.04263.x
PMid:20525006
Edgar R.C., 2004, MUSCLE: multiple sequence alignment with high accuracy and high throughput, Nucleic Acids Res., 32(5): 1792-1797
http://dx.doi.org/10.1093/nar/gkh340
PMid:15034147 PMCid:PMC390337
Finnegan D.J., 1989, Eukaryotic transposable elements and genome evolution, Trends Genet., 5(4): 103-107
http://dx.doi.org/10.1016/0168-9525(89)90039-5
Girard L., and Freeling M., 1999, Regulatory changes as a consequence of transposon insertion, Dev. Genet., 25(4): 291-296
http://dx.doi.org/10.1002/(SICI)1520-6408(1999)25:4<291::AID-DVG2>3.0.CO;2-5
Goulet C., Mageroy M.H., Lam N.B., Floystad A., Tieman D.M., and Klee H.J., 2012, Role of an esterase in flavor volatile variation within the tomato clade, Proc. Natl. Acad. Sci., USA, 109(46): 19009-19014
http://dx.doi.org/10.1073/pnas.1216515109
PMid:23112200 PMCid:PMC3503167
Havecker E.R., Gao X., and Voytas D.F., 2004, The diversity of LTR retrotransposons, Genome Biol., 5(6): 225
http://dx.doi.org/10.1186/gb-2004-5-6-225
PMid:15186483 PMCid:PMC463057
Hua-Van A., Le Rouzic A., Boutin T.S., Filée J., and Capy P., 2011, The struggle for life of the genome's selfish architects, Biol. Direct, 6: 19
http://dx.doi.org/10.1186/1745-6150-6-19
PMid:21414203 PMCid:PMC3072357
International Rice Genome Sequencing Project, 2005, The map-based sequence of the rice genome, Nature, 436(7052): 793-800
http://dx.doi.org/10.1038/nature03895
PMid:16100779
Jiang N., Gao D., Xiao H., and van der Knaap E., 2009, Genome organization of the tomato sun locus and characterization of the unusual retrotransposon rider, Plant J., 60(1): 181-193
http://dx.doi.org/10.1111/j.1365-313X.2009.03946.x
PMid:19508380
Kimura M., and Ohta T., 1972, On the stochastic model for estimation of mutational distance between homologous proteins, J. Mol. Evol., 2(1): 87-90
http://dx.doi.org/10.1007/BF01653945
PMid:4668865
Kumar A., and Bennetzen J.L., 1999, Plant Retrotransposons, Annu. Rev. Genet., 33: 479-532
http://dx.doi.org/10.1146/annurev.genet.33.1.479
PMid:10690416
Ma J., Devos K.M., and Bennetzen J.L., 2004, Analyses of LTR-retrotransposon structures reveal recent and rapid genomic DNA loss in rice, Genome Res., 14(5): 860-869
http://dx.doi.org/10.1101/gr.1466204
PMid:15078861 PMCid:PMC479113
Ma J., and Jackson S.A., 2006, Retrotransposon accumulation and satellite amplification mediated by segmental duplication facilitate centromere expansion in rice, Genome Res., 16(2): 251-259
http://dx.doi.org/10.1101/gr.4583106
PMid:16354755 PMCid:PMC1361721
Marchler-Bauer A., Anderson J.B., Chitsaz F., Derbyshire M.K., DeWeese-Scott C., Fong J.H., Geer L.Y., Geer R.C., Gonzales N.R., Gwadz M., He S., Hurwitz D.I., Jackson J.D., Ke Z., Lanczycki C.J., Liebert C.A., Liu C., Lu F., Lu S., Marchler G.H., Mullokandov M., Song J.S., Tasneem A., Thanki N., Yamashita R.A., Zhang D., Zhang N., and Bryant S.H., 2009, CDD: specific functional annotation with the conserved domain database, Nucleic Acids Res., 37(Database issue): D205- D210
http://dx.doi.org/10.1093/nar/gkn845
PMid:18984618 PMCid:PMC2686570
Marchler-Bauer A., and Bryant S.H., 2004, CD-Search: protein domain annotations on the fly, Nucleic Acids Res., 32(Web Server issue): W327- W331
Marchler-Bauer A., Lu S., Anderson J.B., Chitsaz F., Derbyshire M.K., DeWeese-Scott C., Fong J.H., Geer L.Y., Geer R.C., Gonzales N.R., Gwadz M., Hurwitz D.I., Jackson J.D., Ke Z., Lanczycki C.J., Lu F., Marchler G.H., Mullokandov M., Omelchenko M.V., Robertson C.L., Song J.S., Thanki N., Yamashita R.A., Zhang D., Zhang N., Zheng C., and Bryant S.H., 2011, CDD: a conserved domain database for the functional annotation of proteins, Nucleic Acids Res., 39(Database issue): D225- D229
http://dx.doi.org/10.1093/nar/gkq1189
PMid:21109532 PMCid:PMC3013737
Mccarthy E.M., and Mcdonald J.F., 2003, LTR_STRUC: a novel search and identification program for LTR retrotransposons, Bioinformatics, 19(3): 362-367
http://dx.doi.org/10.1093/bioinformatics/btf878
PMid:12584121
Morgante M., De Paoli E., and Radovic S., 2007, Transposable elements and the plant pan-genomes, Curr. Opin. Plant Biol., 10(2): 149-155
http://dx.doi.org/10.1016/j.pbi.2007.02.001
PMid:17300983
Nesbitt T.C., and Tanksley S.D., 2002, Comparative sequencing in the genus lycopersicon: implications for the evolution of fruit size in the domestication of cultivated tomatoes, Genetics, 162(1): 365-379
PMid:12242247 PMCid:PMC1462239
Ranc N., Mu-os S., Santoni S., and Causse M., 2008, A clarified position for solanum lycopersicum var. cerasiforme in the evolutionary history of tomatoes (solanaceae), BMC Plant Biology, 8: 130
http://dx.doi.org/10.1186/1471-2229-8-130
PMid:19099601 PMCid:PMC2657798
Rogers S.A., and Pauls K.P., 2000, Ty1-copia-like retrotransposons of tomato (Lycopersicon esculentum Mill.), Genome, 43(5): 887-894
http://dx.doi.org/10.1139/g00-056
http://dx.doi.org/10.1139/gen-43-5-887
PMid:11081980
Roman A.C., Benitez D.A., Carvajal-Gonzalez J.M., and Fernandez-Salguero P.M., 2008, Genome-wide B1 retrotransposon binds the transcription factors dioxin receptor and Slug and regulates gene expression in vivo, Proc. Natl. Acad. Sci., USA, 105(5): 1632-1637
http://dx.doi.org/10.1073/pnas.0708366105
PMid:18223155 PMCid:PMC2234196
Sabot F., and Schulman A.H., 2006, Parasitism and the retrotransposon life cycle in plants: a hitchhiker's guide to the genome, Heredity, 97(6): 381-388
http://dx.doi.org/10.1038/sj.hdy.6800903
PMid:16985508
SanMiguel P., Gaut B.S., Tikhonov A., Nakajima Y., and Bennetzen J.L., 1998, The paleontology of intergene retrotransposons of maize, Nat. Genet., 20(1): 43-45
http://dx.doi.org/10.1038/1695
PMid:9731528
SanMiguel P., Tikhonov A., Jin Y.K., Motchoulskaia N., Zakharov D., Melake-Berhan A., Springer P.S., Edwards K.J., Lee M., Avramova Z., and Bennetzen J.L., 1996, Nested retrotransposons in the intergenic regions of the maize genome, Science, 274(5288): 765-768
http://dx.doi.org/10.1126/science.274.5288.765
PMid:8864112
Schnable P.S., Ware D., Fulton R.S., Stein6 J.C., Wei F.S., Pasternak S., Liang C.Z., Zhang J.W., Fulton L., Graves T.A., Minx P., Reily A.D., Courtney L., Kruchowski S.S., Tomlinson C., Strong C., Delehaunty K., Fronick C., Courtney B., Rock S.M., Belter E., Du F.Y., Kim K., Abbott R.M., Cotton M., Levy A., Marchetto P., Ochoa K., Jackson S.M., Gillam B., Chen W.Z., Yan L., Higginbotham J., Cardenas M., Waligorski J., Applebaum E., Phelps L., Falcone J., Kanchi K., Thane T., Scimone A., Thane N., Henke J., Wang T., Ruppert J., Shah N., Rotter K., Hodges J., Ingenthron E., Cordes M., Kohlberg S., Sgro J., Delgado B., Mead K., Chinwalla A., Leonard S., Crouse K., Collura K., Kudrna D., Currie J., He R.F., Angelova A., Rajasekar S., Mueller T., Lomeli R., Scara G., Ko A., Delaney K., Wissotski M., Lopez G., Campos D., Braidotti M., Ashley E., Golser W., Kim H., Lee S., Lin J., Dujmic Z., Kim W., Talag J., Zuccolo A., Fan C.Z., Sebastian A., Kramer M., Spiegel L., Nascimento L., Zutavern T., Miller B., Ambroise C., Muller S., Spooner W., Narechania A., Ren L., Wei S., Kumari S., Faga B., Levy M.J., McMahan L., Van Buren P., Vaughn M.W., Ying K., Yeh C.T., Emrich S.J., Jia Y., Kalyanaraman A., Hsia A.P., Brad Barbazuk W., Baucom R.S., Brutnell T.P., Carpita N.C., Chaparro C., Chia J., Deragon J., Estill J.C., Fu Y., Jeddeloh J.A., Han Y.J., Lee H., Li P.H., Lisch D.R., Liu S.Z., Liu Z.J., Nagel D.H., McCann M.C., SanMiguel P., Myers A.M., Nettleton D., Nguyen J., Penning B.W., Ponnala L., Schneider K.L., Schwartz D.C., Sharma A., Soderlund C., Springer N.M., Sun Q., Wang H., Waterman M., Westerman R., Wolfgruber T.K., Yang L.X., Yu Y., Zhang L.F., Zhou S.G., Zhu Q.H., Bennetzen J.L., Kelly Dawe R., Jiang J.M., Jiang N., Presting G.G., Wessler S.R., Aluru S., Martienssen R.A., Clifton S.W., Richard McCombie W., Wing R.A., and Wilson R.K., 2009, The B73 maize genome: complexity, diversity, and dynamics, Science, 326(5956): 1112-1115
http://dx.doi.org/10.1126/science.1178534
PMid:19965430
Su P.Y., and Brown T.A., 1997, Ty3/gypsy-like retrotransposon sequences in tomato, Plasmid, 38(3): 148-157
http://dx.doi.org/10.1006/plas.1997.1310
PMid:9435017
Tam S.M., Mhiri C., Vogelaar A., Kerkveld M., Pearce S.R., and Grandbastien M.A., 2005, Comparative analyses of genetic diversities within tomato and pepper collections detected by retrotransposon-based SSAP, AFLP and SSR, Theor. Appl. Genet., 110(5): 819-831
http://dx.doi.org/10.1007/s00122-004-1837-z
PMid:15700147
The Tomato Genome Consortium, 2012, The tomato genome sequence provides insights into fleshy fruit evolution, Nature, 485(7400): 635-641
http://dx.doi.org/10.1038/nature11119
PMid:22660326 PMCid:PMC3378239
Tian Z., Rizzon C., Du J., Zhu L., Bennetzen J.L., Jackson S.A., Gaut B.S., and Ma J., 2009, Do genetic recombination and gene density shape the pattern of DNA elimination in rice long terminal repeat retrotransposons? Genome Res., 19(12): 2221-2230
http://dx.doi.org/10.1101/gr.083899.108
PMid:19789376 PMCid:PMC2792168
Wicker T., Sabot F., Hua-Van A., Bennetzen J.L., Capy P., Chalhoub B., Flavell A., Leroy P., Morgante M., Panaud O., Paux E., SanMiguel P., and Schulman A.H., 2007, A unified classification system for eukaryotic transposable elements, Nat. Rev. Genet., 8(12): 973-982
http://dx.doi.org/10.1038/nrg2165
PMid:17984973
Xiao H., Jiang N., Schaffner E., Stockinger E.J., and van der Knaap E., 2008, A retrotransposon-mediated gene duplication underlies morphological variation of tomato fruit, Science, 319(5869): 1527-1530
http://dx.doi.org/10.1126/science.1153040
PMid:18339939
Xiong Y., and Eickbush T.H., 1990, Origin and evolution of retroelements based upon their reverse transcriptase sequences, EMBO J., 9(10): 3353-3362
PMid:1698615 PMCid:PMC552073
Ya-ez M., Verdugo I., Rodríguez M., Prat S., Ruiz-Lara S., 1998, Highly heterogeneous families of Ty1/copia retrotransposons in the Lycopersicon chilense genome, Gene, 222(2): 223-228
http://dx.doi.org/10.1016/S0378-1119(98)00486-7